Neurobiologie et dynamique de la perception bistable de stimuli ambigus
Le comportement efficace et adapté requiert une analyse rapide et fiable de l’information sensorielle. Confrontés à des scènes naturelles, les systèmes sensoriels doivent analyser un flot d’information complexe voire embrouillé, et souvent ambigu. Une organisation perceptuelle stable et fiable de la stimulation sensorielle doit pourtant être élaborée rapidement afin de guider le comportement. Les mécanismes neuronaux responsables de cette organisation perceptuelle peuvent être mis à jour, notamment chez l’homme, par l’utilisation de situations intrinsèquement ambiguës. On peut retenir au moins trois phénomènes remarquables lors de la perception de stimuli ambigus.
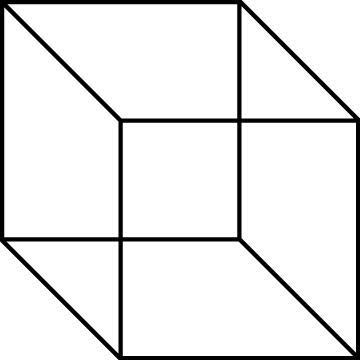
1 – La perception, elle, n’est pas ambiguë. La perception d’un cube de Necker, par exemple, qui utilise l’ambiguïté de la projection en deux dimensions d’une image tridimensionnelle, n’est pas incertaine. Il est tout d’abord très rare de percevoir le dessin comme celui d’un objet plat – cela montre la mise en œuvre automatique de mécanismes actifs de reconstruction tridimensionnelle (on a donc un outil pour étudier la logique de l’organisation perceptuelle). Ensuite, bien que la face du cube perçue ‘en avant’ puisse être celle du haut ou celle du bas, à chaque instant une seule de ces interprétations est accessible à la conscience.
2 – Le corollaire du point précèdent est qu’il est ainsi possible de dissocier la perception consciente des caractéristiques de la stimulation physique : un stimulus identique peut évoquer des percepts différents.
3 – La perception est bistable : sans que le stimulus ne change, l’interprétation change spontanément au cours du temps. L’origine de cette dynamique perceptuelle est encore mal comprise, mais il est probable qu’elle reflète des caractéristiques intrinsèques de l’accès à la conscience de l’interprétation perceptuelle. L’intérêt de la perception bistable pour l’étude de la neurobiologie de la conscience provient du fait que la dynamique de la bistabilité semble obéir à des règles strictes, observables pour des stimuli visuels très différents, ainsi que pour des stimuli auditifs ambigus. On peut dès lors supposer que la dynamique de la bistabilité reflète des principes fondamentaux du traitement cortical permettant la construction de perceptions conscientes.
Publications
Hupé, J. M., Lamirel, C., & Lorenceau, J. (2009).
Pupil dynamics during bistable motion perception. Journal of Vision,
9(7:10), 1-19. [Article]
Hupé, J. M., Lamirel, C., & Lorenceau, J. (2008). Pupil dilation does not predict subsequent stability in perceptual rivalry. Proc Natl Acad Sci U S A, 105(28), E43; author reply E44. [Article]
Hupé, J.-M., Joffo, L.-M., & Pressnitzer, D. (2008). Bistability for audiovisual stimuli: Perceptual decision is modality specific. Journal of Vision, 8(7:1), 1-15. [Article]
Pressnitzer, D., &
Hupé, J. M. (2006). Temporal dynamics of auditory and visual
bistability reveal common principles of perceptual organization.
Current Biology, 16(13), 1351-1357.
pdf
Supplementary Material (pdf)
demo
Rubin, N., & Hupé, J.-M. (2005). Dynamics of
perceptual bi-stability: plaids and binocular rivalry compared. In A.
Alais & R. Blake (Eds.), Binocular Rivalry (Chapter 8). Cambridge:
MIT Press.
Preprint pdf
Hupé, JM & Rubin, N. (2003) The dynamics of
bi-stable alternation in ambiguous motion displays: a fresh look at
plaids. Vision Res 43: 531 - 548.
Preprint pdf
Information Supplémentaire, Démo
Hupé, JM, & Rubin, N. (2001) Transparent motion
is always more likely for plaids moving along oblique directions than
for plaids moving along cardinal directions. IOVS, 42 (Suppl.), 736.
(voir la démo présentée à l’ARVO en 2001)
Hupé, JM & Rubin, N. (2004) The Oblique Plaid Effect. (Preprint pdf)
Hupé, JM, & Rubin, N. (2000) Perceived motion
transparency can override luminance / color cues which are inconsistent
with transparency. IOVS, 41 (Suppl.), 721.
(voir la démo présentée à l’ARVO en 2000)
 Retour à la page d'accueil
Retour à la page d'accueil